Fopius Wharton, 1987
Type locality of type species: Cameroon, Nkolbisson; holotype female in US National Museum.
First described as a subgenus, then elevated to generic rank by van Achterberg and Maeto (1990).
Current status: valid genus.
Prior to 1987, most of the species now placed in the genus Fopius were placed either in Opius or Biosteres, and sometimes in Diachasma or Parasteres. Wharton (1987) initially proposed the name Fopius as a subgenus of Rhynchosteres. Van Achterberg and Maeto (1990) elevated Fopius to generic status, where it currently remains. Wharton (1999) partially revised the genus, and included a tabular key to species.
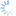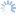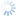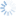
Fopius was originally described as a subgenus of Rhynchosteres (Wharton 1987), despite the fact that the type species of Fopius lacked the highly modified, tubular clypeus originally used to characterize Rhynchosteres. Wharton’s (1987) placement of Fopius in Rhynchosteres was based on a study of several Afrotropical species that had the midventral margin of the clypeus variously thickened and protruding, suggesting a morphocline leading to the more elaborate modifications found in Rhynchosteres sensu stricto. Van Achterberg and Maeto (1990), dissatisfied with the broadening of the concept of Rhynchosteres that this entailed, removed Fopius from Rhynchosteres and accorded it separate generic rank. The Fopius desideratus species group is particularly critical for an understanding of the relationship between Fopius and Rhynchosteres. Althought the mandible lacks the modifications found in most species of Rhynchosteres, the midventral portion of the clypeus in members of the Fopius desideratus species group is at least slightly thickened and weakly protruding. Additionally, the relative length of the first two flagellomeres in the Fopius desideratus species group is as in Rhynchosteres, the petiole is similarly structured (flatter and narrower than in more typical species of Fopius such as Fopius silvestrii), and most significantly, the ovipositor sheath shows an identical pattern of setal reduction.
Chinajariyawong et al. provide an extensive list of opiine species and their tephritid hosts collected in Thailand and Malaysia during surveys conducted between 1986 and 1994 (Chinajariyawong et al. 2000). The specimens were primarily collected from various Bactrocera spp. and include the following Fopus species: arisanus (Sonan), deeralensis (Fullaway), persulcatus (Silvestri), skinneri (Fullaway), and vandenboschi (Fullaway) with arisanus and vandenboschi being the most commonly collected species of Fopius.
Fopius persulcatus species group
The Fopius persulcatus species group is characterized largely by striate sculpture on the second metasomal tergum. The frons is also densely covered with large punctures, these sometimes coalescing to form rugulose or rugulo-striate sculpture. This species group consists of Fopius alternatae (Tobias) and Fopius myolejae (Tobias) from Russia; and the Indo-Pacific species Fopius arisanus (Sonan), Fopius carpomyiae (Silvestri), Fopius persulcatus (Silvestri), Fopius skinneri (Fullaway), and Fopius vandenboschi (Fullaway). The species are very similar to one another, but differ primarily in coloration, length of ovipositor, and configuration of the ovipositor tip. Following their successful introduction to Hawaii during the biological control program against oriental fruit fly (Clausen et al. 1965), Fopius arisanus and Fopius vandenboschi were intensively studied, and much is now known about their biology (with most of the early literature on arisanus published under the name Opius oophilus Fullaway). Other than the original host records, however, little is known about the other species. Data on persulcatus are particularly problematic because of widespread confusion regarding the identity of this species during the Hawaiian oriental fruit fly program, and the subsequent description of several subspecies (Fischer 1963). Though the best known species are parasitoids of tephritids in the dacine genus Bactrocera Macquart in their native range, other species have been reared from trypetine tephritids in the genera Carpomya Costa, Myoleja Rondani, and Rhagoletis Loew. Based on the similarities in the shape of the ovipositor tip, these species preferentially attack either the egg or early instar larva of their host (though all eventually emerge from the puparium). This biology, however, has only been confirmed for arisanus (attacking eggs) and vandenboschi (attacking primarily first instars). The species of this group are known from Pakistan and India east through Indonesia and north through Taiwan, Japan, and far eastern Russia.
Fopius desideratus species group.
Members of this species group that have been reared from fruit-infesting tephritids include Fopius bevisi (Brues), Fopius desideratus (Bridwell), Fopius niger (Szépligeti), Fopius ottotomoanus (Fullaway) and Fopius okekai Kimani-Njogu and Wharton. As noted above, this group is characterized by the reduced setae on the ovipositor sheath, slight thickening medially along the ventral margin of the clypeus, and a longer, somewhat flatter petiole than is found in members of the Fopius silvestrii group (discussed below). The punctation on the frons tends to be weaker, with transverse striae usually clearly present medially. The species of the Fopius desideratus species group are very similar to one another, differing primarily in color. Members of this species group have been collected from Cameroon and Nigeria east to Kenya and Tanzania as well as South Africa and Madagascar, and undoubtedly occur throughout subsaharan Africa. Most but not all of the specimens reared to date have been from Dacus infesting Cucurbitaceae. The few remaining records were from ceratitidine tephritids on various hosts. All members of this species group have large, subapical nodes on the ovipositor, suggesting that they may attack late instar larvae of their hosts.
Fopius silvestrii species group.
This group is currently defined largely by the absence of features that define the two species groups already mentioned: the second metasomal tergum is unsculptured and the setae on the ovipositor sheath are not reduced. Both Fopius silvestrii (Wharton) and Fopius ceratitivorus Wharton have been reared from medfly, Ceratitis capitata, infesting coffee, and Fopius silvestrii has also been reared from Dacus bivittatus (Bigot) infesting squash (Steck et al. 1986, Wharton 1987). The two other members of this species group have not been reared. Members of the Fopius silvestrii species group have much the same distribution pattern as those of the Fopius desideratus species group, and are differentiated from one another largely by color (Fopius silvestrii is dark and Fopius ceratitivorus is pale) and ovipositor length. Fopius ceratitivorus is known only from Kenya and extreme northeastern South Africa; Fopius silvestrii has been collected in West Africa and western Kenya.
Other species groups
The remaining three species, Fopius caudatus (Szépligeti), Fopius deeralensis (Fullaway), and Fopius schlingeri Wharton, do not readily cluster into distinctive species groups. Fopius caudatus is readily separated from all other species of Fopius by the distinctive band of setae and punctures on the frons extending between the eye and ocelli. Fopius caudatus resembles members of the Fopius desideratus species group in the morphology of the clypeus and petiole, but has a distinctly different ovipositor (strongly narrowed, suggesting oviposition in the host egg) as well as several features unusual for members of the genus Fopius (Wharton 1997). The commonly collected Fopius caudatus also superficially resembles Fopius silvestrii, and both have been reared from ceratitidines in coffee samples. Fopius caudatus has thus far been reared exclusively from ceratitidines (Steck et al. 1986, Wharton et al. 2000), and is known from tropical regions of both eastern and western Africa.
Both Fopius deeralensis and Fopius schlingeri are from Queensland, where they have been reared from various species of Bactrocera in a variety of host fruits (Clausen et al. 1965, Wharton 1997). Fopius schlingeri, for example, attacks at least four species of Bactrocera developing in the fruit of at least three plant families. Fopius deeralensis differs from Fopius schlingeri in having the ventral margin of the clypeus distinctly pointed midventrally, a distinct subapical ridge on the ovipositor, and the second metasomal tergum is usually weakly striate, at least basally. There are slight differences among species of Fopius in the exposure of the labrum and Fopius schlingeri provides a good example of a species with a partially exposed labrum. The ovipositor in Fopius schlingeri is virtually identical in form to that of Fopius arisanus, strongly suggesting a biology similar to the latter in which the parasitoid oviposits into the egg of its host. Field observations kindly supplied by Greg Quimio during his PhD research at the University of Queensland appear to confirm this. These two Australian species differ in only minor ways from members of the Fopius persulcatus group, and may need to be included there after further study.












Unless you are already on this page, considerably more information on species groups and use in biological control can be found on the Fopius page (and associated links) of the Paroffit website page: http://mx.speciesfile.org/projects/8/public/public_content/show/8319?content_template_id=88
