Dirhinus giffardii Silvestri
Remarks
For additional information, see the Dirhinus page.
Description
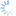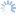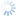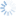
Like all other members of the genus Dirhinus, D. giffardii can be recognized by the antennae, which are inserted in a deep concavity formed by two ridges extending out from the face (Figs. 1-3). In dorsal view, these appear as two horn-like protruberances.



Diagnosis and Relationships
Relationships largely unknown; the species is in need of revision. Boucek and Narendran (1981) revised the species of Dirhinus from India, and synonymized several species under the name Dirhinus anthracia Walker based in part on the fact that previous authors treating regional faunas had overlooked the cosmopolitan distribution of this species. There is a good possibility that at least some of the material previously determined as Dirhinus giffardii is in fact D. anthracia.
Distribution
Originally described from Nigeria (Silvestri 1914).
Subsequently introduced to Hawaii (Purcell 1998) and Israel (Rivnay 1968).
There is also a report of breeding this species on dacines in Pakistan (Ahmad et al. 1975).
Subsequently introduced to Hawaii (Purcell 1998) and Israel (Rivnay 1968).
There is also a report of breeding this species on dacines in Pakistan (Ahmad et al. 1975).
Distribution
No referenced distribution records have been added to the database for this OTU.
Biology / Hosts
Originally reared from Ceratitis anonae Graham in Nigeria and propagated on medfly, Ceratitis capitata (Silvestri 1914). Dirhinus giffardii is a generalist parasitoid known to attack a wide variety of other fruit-infesting tephritids including Anastrepha obliqua (Macquart), A. suspensa (Loew), Ceratitis rosa Karsch, Bactrocera cucurbitae (Coquillett), B. demmerezi (Bezzi), B. dorsalis (Hendel), B. oleae (Rossi), B. passiflorae (Froggatt), Neoceratitis cyanescens (Bezzi), B. tryoni (Froggatt), and Toxotrypana curvicauda Gerstaecker (Noyes’ Universal Chalcidoidea database, but updated here for nomenclature of hosts).
Biology and Behavior
An ectoparasitoid of tephritid fruit fly pupae, female Dirhinus giffardii pierce the puparial wall and oviposit onto the host pupa (Silvestri 1914). The host continues to develop until the parasitoid larva hatches, at which time the host is paralyzed and consumed by the larva. Wang and Messing (2004) provide evidence that in laboratory settings D. giffardii females prefer to attack larger host species when given a choice, resulting in larger parasitoid offspring. The fitness gained by the offspring in foraging efficiency due to larger size did not appear to come at the cost of development time or survival. See also Wang and Messing (2004a).
Biology - Host Range Testing
Dirhinus giffardii has also been reported to attack other parasitoids, including Fopius vandenboschi (Dresner 1954) creating a concern about the potential impact of introduced D. giffardii on nontarget species. In additional laboratory studies, Wang and Messing (2004), studied the ability of Dirhinus giffardii to attack parasitoids used for biological control in Hawaii, including Fopius arisanus (Sonan), Diachasmimorpha longicaudata (Ashmead), Diachasmimorpha tryoni (Cameron), and Psyttalia incisi (Silvestri). They found that D. giffardii readily hyperparasitizes Ceratitis capitata previously parasitized by any of the 4 parasitoid species. However, when given a choice D. gifffardii females preferred to oviposit in unparasitized fly pupae. This preference was correlated with greater overall fitness experienced by the parasitoid offspring reared on the primary host.
Biological Control
Dirhinus giffardii (Silvestri) was brought from Africa to Hawaii by Silvestri (1914). In Hawaii, it was propagated, released, and became established on medfly, Ceratitis capitata. Later (1971), this species was introduced as a biological control agent in Bolivia against medfly (Bennett and Squire 1972). It has also been released in China (Chitung Chen, personal communication).
